Содержание
- Лекция: Репарация генетического материала, её биологическое значение, механизм и системы
- Повреждения ДНК спонтанные и индуцированные. Процессы репарации ДНК и их возможные последствия. Мутации. Роль мутаций в эволюции и возникновении наследуемых заболеваний. Понятие о генной терапии.
- Репарация (биология)
- Лекция: Репарация генетического материала, её биологическое значение, механизм и системы
- Коррекция нуклеотидной последовательности при репликации
- Репарация сразу после репликации
- Репарационные механизмы при повреждении ДНК
- Устранение ошибок ДНК-полимеразой
- Репарационные механизмы
- 266. Репарация днк. Виды репарации.
- Репарация ДНК
Лекция: Репарация генетического материала, её биологическое значение, механизм и системы
Важное значение для ограничения неблагоприятных последствий генных мутаций имеют естественные антимутационные барьеры.Одним из них является парность хромосом в диплоидных наборах хромосом эукариот, которая препятствует проявлению рецессивных мутаций у гетерозиготных особей. Главным антимутационным барьером рассматривается выработавшая в процессе эволюции способность к репарации наследственного материала. Её сущность — в устранении из наследственного материала клетки изменённого участка.
Различают 3 системы репарации генетического материала:эксцизионная репарация (репарация путём «вырезания»), фоторепарация и пострепликативная репарация.
Механизм эксцизионной репарации заключается в ферментативном разрушении изменённого участка молекулы ДНК с последующим восстановлением на этом отрезке нормальной последовательности нуклеотидов. Такой механизм включает следующие этапы (рис. 81): а) разрыв спирали ДНК у места повреждения при участии эндонуклеаз; б) удаление повреждённого участка с запасом в обе стороны с помощью эндонуклеаз; в) синтез при участии ДНК-полимеразы на месте дефекта нормального участка ДНК;
г) «сшивание» последнего с образовававшимися концами спирали ДНК при помощи фермента ДНК-лигазы (восстановление непрерывности ДНК).
Например, под действием УФ-лучей у человека нарушается комплементарность пар нуклеотидов в двойной спирали ДНК (появляются пары Т-Т, Ц-Ц и т.п.). Они устраняются вышеописанным способом. Однако у различных индивидуумов наблюдаются генетические различия в активности репаративных ферментов и надёжности функционирования механизма ферментативного разрушения изменённого участка молекулы ДНК в целом. У ряда людей наблюдается изменение ДНК и, как следствие, возникновение заболевания «пигментная ксеродерма».
В клетках эукариот обнаружены два вида репарации «путём вырезания»:1) более продолжительная репарация (длительность процесса — от 1 до 24 часов), восстанавливающая большой фрагмент ДНК (около 100 нуклеотидов); 2) быстродействующая репарация (продолжается от 5 минут до 2 часов), восстанавливающая 3-4 нуклеотида.
Пострепликативная репарация«включается» тогда, когда эксцизионная репарация «не справляется» с устранением всех повреждений, возникших в ДНК до её репликации. При репликации во второй спирали ДНК возникают бреши — однонитевые пробелы, соответствующие изменённым нуклеотидам первой спирали. Бреши заполняются участками цепи с нормальной последовательностью нуклеотидов уже в ходе пострепликативной репарации при участии ДНК-полимеразы.
Фоторепарация заключается в расщеплении ферментом (дезоксирибопиримидинфотолиазой), активируемым видимым светом, циклобутановых димеров, возникающих в ДНК под действием ультрафиолетового излучения.
Механизмам репарации свойственны нарушения и «сбои», которые приводят к повышению чистоты мутаций. Известны специфические мутации, блокирующие механизмы репарации и вызывающие наследственные заболевания (пигментная ксеродерма и др.).
Биологическое значение репарации ДНК заключается в резком снижении частоты мутаций, большинство которых оказываются летальными и полулетальными или же снижающими жизнеспособность организмов, вызывающими аномалии и обусловливающими тератогенез. Благодаря репарации ДНК повышается устойчивость генотипа организма к повреждающим агентам (мутагенам).
Повреждения ДНК спонтанные и индуцированные. Процессы репарации ДНК и их возможные последствия. Мутации. Роль мутаций в эволюции и возникновении наследуемых заболеваний. Понятие о генной терапии.
Повреждение структуры ДНК
Ошибки при репликации, то есть встраивание некомплементарных оснований тут же исправляются репаративными системами
В репаративную систему входят:
— белки-ферменты узнающие ошибку
— белки- ферменты разрезающие в этом месте цепочку
— ДНК-полимеразы достраивающие нить
— ДНК-лигазы (сшивающие нить) завершающие репарацию
Нарушения комплементарности цепей ДНК могут происходить спонтанно, т.е. без участия каких-либо повреждающих факторов, например в результате ошибок репликации, дезаминирования нуклеотидов, депуринизации.
Индуцированные повреждения ДНК между репликациями могут возникать под действием УФ, рентгеновского излучения, химических агентов и свободных радикалов /При таких повреждениях репаративные системы могут быть не состоятельны, изменения ДНК сохраняются – образуются мутации/
Мутации (от лат. mutatio — изменение), внезапные (скачкообразные) естественные или вызванные искусственно наследуемые изменения генетического материала (генома), приводящие к изменению тех или иных признаков организма.
/мутации являются материалом для естественного отбора// мутации в ДНК половых клеток вызывают наследственные заболевания/
ГЕННАЯ ТЕРАПИЯ- это лечение болезней путем введения пациенту здоровых ГЕНОВ вместо недостающих или поврежденных. Первый раз человека подвергли такому лечению в США в 1990 г./ Здоровый ген вводят в какой-нибудь вирус (обычно переносчик легко поддающейся лечению инфекции) так, чтобы он был непосредственно нацелен на поврежденные клетки./
Регуляция клеточного цикла и репликации. Роль циклинов и белка Р53.
Регуляция репликации
Длительность фаз клеточного цикла различна у различных типов клеток (более 200 видов) G1-G0- фаза покоя, при этом некоторые клетки не могут перейти в S-фазу (репликации) (нейроны, миоциты)
Основным фактором, который включает, замедляет, восстанавливает репликацию в S-фазу клеточного цикла, а также регулирует прохождение клеток по другим фазам клеточного цикла является повреждение ДНК и мутации.
В каждой фазе клеточного цикла осуществляется контроль структуры ДНК и если в ней находятся ошибки – прохождение клетки по клеточному циклу затормаживается, а если ошибка не исправляется – цикл останавливается.
Главную роль в регуляции клеточного цикла и репликации играет белок р53.
/синтезируется во многих клетках с гена р53 ^является «молекулярным полицейским», который улавливает, узнает ошибки в первичной структуре ДНК/
При обнаружении ошибок, белок р53 предпринимает следующие действия:
— активирует ферменты репаративной системы
— разрушает циклины, отдаляя время репликации
— если ошибки не устранены – запускается апопптоз (в любой из фаз клеточного цикла) р53 запускает синтез каспаз и ДНКаз, которые разрушают белки и ДНК, что приводит к смерти клетки (физиологической)
/р53 активирует биосинтез белков семейства bax (активаторы каспаз)/
р53 активирует биосинтез белка fаs/аро1 – трансмембранный гликопротеид, который появляется в мембране клетки при запускании апоптоза (в норме его нет) и выполняет рецепторную роль для макрофагов, с целью фагоцитоза
/р53 ингибирует биосинтез белка bcl-2 (ингибитор каспаз)./
Но сам ген кодирующий белок р53 может мутировать:
— утрачивается контроль за структурой ДНК
— апоптоз не включается
— происходит без контрольное деление клеток с поврежденной ДНК
— такие нарушения лежат в основе развития рака
9.Апоптоз. Физиологическая роль, механизмы развития. Роль белка Р53, последствия мутаций в гене р53. Биохимические основы противоопухолевой терапии, значение лабораторного определения маркеров апоптоза.
Апоптоз – процесс естественной смерти клетки, морфологически проявляется прогрессирующей фрагментацией клеточных компонентов, включая ДНК
Происходит гидролитический распад белков под действием протеаз, называемых каспазами и распад ДНК с помощью ДНКаз [Эти ферменты не находятся в лизосомах, в отличии процессов, происходящих при некрозе, где работают лизосомальные ферменты]
Апоптоз –физиологический процесс, активирующийся не только у мутагенных клеток, но также активируется у высокоспециализированных клеток
/Например у клеток, формирующих иммунный ответ организма, при воспалении, после устранения патогена, у лейкоцитов активируется аппоптоз./
Но сам ген кодирующий белок р53 может мутировать:
— утрачивается контроль за структурой ДНК
— апоптоз не включается
— происходит без контрольное деление клеток с поврежденной ДНК
— такие нарушения лежат в основе развития рака
Стратегия противоопухолевой терапии – активация апоптоза опухолевых клеток. Это достигается путем радиоактивного облучения, химического воздействия (химиотерапия)
В настоящее время в клинико-диагностических лабораториях можно оценить экспрессию генов в опухолевых клетках методом люминесцентной микроскопии, с целью оценки эффективности противоопухолевой терапии :
— р53,
— fаs/аро1,
— bcl-2
Если терапия корректна, то в опухолевых клетках повышается экспрессия генов р53 и fаs/аро1 и супрессируется ген bcl-2
Транскрипция. Основные элементы транскриптона.
Репарация (биология)
Компоненты, необходимые для транскриции. Механизм и биологическое значение транскрипции.
Транскрипция – перенос генетической информации от ДНК к мРНК /переписывание генетической информации в виде последовательности дезокрибонуклеотидов в последовательность рибонуклеотидов/
Биологическое значение – синтез мРНК, которая выполняет роль матрицы для синтеза полипептида (белка) [При транскрипции переписывается не весь геном, а лишь некоторый участок ДНК, называемый геном]
Транскриптон — участок матричной ДНК, с которой происходит процесс переписывания, то есть транскрипция /Состоит из нескольких участков
Промотор – участок ДНК, к которому присоединяется фермент РНК-полимераза
Оператор — участок ДНК, к которому присоединяются различные белки, регулирующие скорость транскрипции
Терминатор — стоп сигнал, завершающий транскрипцию./
3 этапа транскрипции:
— инициация
— элонгация
— терминация
Инициация транскрипции /Распознается большая бороздка ДНК =РНК-полимераза находит промотор в ДНК и взаимодействует с ним =ДНК на этом участке начинает плавиться (расплетаются нити ДНК) = Промотор содержит пары А-Т, поэтому плавится достаточно легко/
Элонгация транскрипции (синтез РНК) /для элонгации необходимо:рибонуклеотиды, служащие субстратом и источником энергии для этого процесса;фермент ДНК-зависимая –РНК- полимераза;в процессе элонгации мононуклеотиды во вновь синтезированной цепи РНК строго комплементарны мононуклеотидам матричной ДНК/
Терминация транскрипции /достижение РНК-полимеразы стоп-сигнала и отсоединение синтезированной РНК/
Созревание и процессинг мРНК
— При синтезе мРНК транскрибируется ДНК, в которую входят участки информационные и ненесущие информации, причем их до 90%
— После транскрипции участки РНК ненесущие информацию, называемые интронами вырезаются
— Остаются только информативные участки РНК
— Из ядра в цитоплазму выходит созревшая РНК.
Генетический код. Свойства генетического кода, биологическое значение.
Генетический код – запись информации о первичной структуре белка (последовательности АК) в форме последовательности рибонуклеотидов в
м-РНК.
Свойства генетического кода
1)Специфичность – каждой аминокислоте соответствует триплет нуклеотидов
2)Триплетность – кодон состоит из 3-х нуклеотидов
3)Вырожденность – одной аминокислоте соответствует несколько кодонов
4)Непрерывность – между кодонами нет нуклеотидов, разделяющих их
5)Неперекрываемость — каждый нуклеотид входит в состав лишь одного кодона
6)Универсальность – у всех живых организмов одни и те же кодоны несут информацию об одних и тех же аминокислотах
7)Коллинеарность – соответствие линейной последовательности нуклеотидов в м-РНК линейной последовательности аминокислот в белке
Репарация ДНК — это ее починка, т. е. исправление ошибок, возникающих в структуре молекулы. Слово «репарация» происходит от английского «repair», переводимого как «ремонт», «починка» и т. п.
Под ошибками в структуре ДНК, которые могут быть репарированы, чаще всего понимают нарушение последовательности нуклеотидов — структурных единиц, из которых состоит каждая цепь ДНК. Молекула ДНК состоит из двух цепей-нитей, комплементарных друг другу. Это значит, что если повреждения возникают в одной из цепей, то по второй неповрежденной можно восстановить испорченный участок первой. Кроме этого, в клетках эукариот каждая хромосома имеет гомологичную, т. е. содержащую тот же набор генов (но не аллелей). В крайнем случае, когда поврежден участок на обеих нитях молекулы, он может копироваться с гомологичной хромосомы. Также после S-фазы клеточного цикла, когда произошла репликация (самокопирование), каждая хромосома состоит из двух двухцепочечных идентичных друг другу хроматид, т. е. по-сути из двух идентичных молекул ДНК.
Лекция: Репарация генетического материала, её биологическое значение, механизм и системы
Это также может быть использовано для восстановления исходной структуры поврежденной молекулы.
В процессе эволюции появилось много различных клеточных молекулярных механизмов, ответственных за репарацию ДНК. В основном это различные ферменты и их комплексы. Часть из них участвует также в репликации. Особо опасны повреждения генов, которые кодирую такие ферменты. Это приводит к утрате того или иного репарационного механизма. В этом случае в клетках происходит более быстрое накопление повреждений и мутаций. Нередко это служит причиной возникновения бесконтрольно делящихся клеток, т. е. появления опухолей.
С другой стороны, если повреждения ДНК особенно сильны, то в клетках включается механизм самоуничтожения (апоптоза). Таким образом к делению такие клетки не допускаются, а значит следующее поколение не будет содержать значительные повреждения ДНК.
Ошибки в структуре ДНК могут возникать на различных этапах ее существования (во время синтеза, в пред- и постсинтетические периоды), по разным причинам (случайно, под действием химически активных веществ, радиации и др.). Также изменения бывают разными (потеря химической группы нуклеотида или присоединение дополнительной, замена нуклеотида на другой, установление химической связи между двумя соседними нуклеотидами, разрыв цепи, потеря участка и др.). В связи с таким разнообразием существует трудность классификации репарационных механизмов. Часто их делят на те, которые происходят во время репликации, сразу после нее и в течение остального жизненного цикла клетки. Ниже перечислены наиболее изученные причины изменения структуры ДНК и способы репарации.
Следует иметь в виду, что не все ошибки исправляются, относительно мелкие и не критичные могут передаваться следующему поколению клеток и организмов. Их нельзя назвать повреждениями, скорее — мутациями. Большинство мутаций вредны, однако те, что нейтральны или полезны в данных условиях окружающей среды, служат материалом для эволюции. Таким образом несовершенство механизмов репарации ДНК обеспечило разнообразие жизни на нашей планете.
Коррекция нуклеотидной последовательности при репликации
ДНК-полимеразы выполняют основную работу при репликации ДНК, присоединяя нуклеотид за нуклеотидом к новой цепи. Помимо основной функции, многие полимеразы способны удалять неправильно присоединенный последний нуклеотид, т. е. не комплементарный нуклеотиду матричной цепи.
Химическая структура нуклеотидов может несколько модифицироваться. При этом они начинают соединяться водородными связями не со своими комплементарными напарниками. Так, например, цитозин должен связываться с гуанином. Но его измененная форма устанавливает водородные связи с аденином, с которым должен был связаться тимин.
При синтезе новой нити ДНК очередной нуклеотид сначала связывается водородными связями с комплементарным основанием матрицы. После этого полимераза связывает его с концом растущей цепи ковалентной связью.
Однако, если это был модифицированный нуклеотид, который неправомерно связался с комплементарным основанием материнской цепи, то он обычно быстро возвращается в свою исходную форму и становится некомплементарным. Водородные связи разрываются, и получается, что конец новой цепи имеет свободно висящий нуклеотид, ковалентно связанный с синтезируемой цепью.
ДНК-полимераза в данном случае не может присоединить следующий нуклеотид, и ей ничего не остается, как только удалить этот ошибочный нуклеотид.
Если же водородные связи не разорвались, то за ошибочным нуклеотидом цепь продолжит нарастать далее, а точечная мутация сохранится. Она может быть устранена уже после репликации.
Репарация сразу после репликации
После того, как новая нить ДНК была синтезирована, определенные комплексы ферментов распознают неправильно спаренные основания. При этом существует проблема определения новой и старой цепей молекулы ДНК. Новая отличается отсутствием метилированных оснований и у эукариот наличием временных разрывов. По этим признакам ферментные комплексы идентифицируют именно вновь синтезированную цепь. Таким образом в некомплементарных парах оснований «ошибкой» считается нуклеотид новой цепи.
Как только ошибка найдена, другие ферменты вырезают целый участок ДНК, содержащий неправоменое основание, а не только один нуклеотид. После этого полимераза заново строит этот участок, а лигаза сшивает его с остальной цепью. Этот механизм, когда вырезается и вновь синтезируется участок ДНК, называется эксцизионной репарацией (от слова excision — отрезание, вырезание), он достаточно универсален и используется во многих случаях репарации, а не только при «проверке» ДНК сразу после репликации.
Репарационные механизмы при повреждении ДНК
ДНК организма может изменяться не только из-за ошибок во время репликации. Клетка живет, подвергается воздействию неблагоприятных внешних факторов, ее внутренняя биохимическая среда может изменяться, провоцируя пагубные для ДНК реакции. В результате генетический материал так или иначе повреждается. В зависимости от типа повреждения, его масштаба включаются различные репарационные механизмы, привлекающие несколько различающиеся наборы ферментативных комплексов.
1. Существуют ферменты, отменяющие изменения нуклеотидов на месте без удаления участков ДНК. Другими словами, если в цепи был нуклеотид, содержащий основание гуанин (Г), который в результате химической реакции присоединил метил-группу и превратился в метил-гуанин, то фермент превратит его обратно в гуанин. В основном подобная репарация ДНК касается присоединения-отсоединения определенных групп атомов.
2. В случае утраты пуриновых оснований может протекать эксцизионная репарация. В случае дезаминирования и некоторых других структурных изменений оснований, ферменты гликозилазы вырезают только поврежденное основание нуклеотида. И только после этого протекает стандартная эксцизионная репарация.
3. Вырезается участок и при образовании димеров, когда два соседних нуклеотида соединяются между собой. Обычно такие реакции протекают в результате воздействия ультрафиолетовых лучей. Образование димера провоцирует расхождение комплементарных нитей ДНК в этом и близлежащих участках. Образуется пузырь, который распознается ферментами. Далее запускается эксцизионная репарация.
4. Бывают столь сильные повреждения молекул ДНК, когда структура обеих ее цепей нарушается в одном и том же месте. При этом уже нельзя согласно принципу комплементарности восстановить одну цепь по другой. Одним из примеров подобного повреждения может является разрыв молекулы ДНК на две части, например, при действии сильного радиоактивного облучения.
В случае повреждения обеих нитей молекулы ДНК на помощь может прийти рекомбинативная репарация, когда вместо поврежденного участка вставляется участок с гомологичной хромосомы или сестринской хроматиды. В случае разрыва также существуют ферменты, способные обратно присоединять оторванный кусок ДНК. Однако при этом часть нуклеотидов может теряться, что в свою очередь может привести к серьезным мутациям.
Рекомбинативная репарация в пресинтетический период клеточного цикла может протекать только между гомологичными хромосомами, т. к. каждая хромосома в этот период состоит только из одной хроматиды. В постсинтетический период, когда хромосомы состоят из двух идентичных хроматид, участок может заимствоваться с сестринской хроматиды.
Следует подчеркнуть, что у сестринских хроматид набор аллелей исходно идентичен (если не было кроссинговера). У гомологичных хромосом — нет. Таким образом, настоящая рекомбинация с точки зрения генетики протекает только в случае обмена между гомологичными хромосомами. Хотя здесь в обоих случаях мы говорим о рекомбинации.
Рассмотрим такой пример. Допустим в ДНК возник тиминовый димер, который не был репарирован до репликации. В процессе репликации цепи исходной молекулы ДНК расходятся и на каждой строится новая комплементарная цепь. На той матричной цепи, которая содержит димер тимина, в этом участке не может быть построен участок новой цепи. В этом месте просто отсутствует нормальный шаблон. В дочерней нити появляется брешь, а в материнской остается димер. Т. е. данная молекула ДНК «не знает», какова правильная нуклеотидная последовательность участка.
Единственный выход в данном случае – позаимствовать кусок ДНК с другой хроматиды. Он переносится с одной из ее цепей. Образовавшаяся здесь брешь застраивается по шаблону комплементарной цепи. Перенесенный участок на поврежденной молекуле застраивает брешь дочерней цепи, материнская так и продолжит содержать димер, который может быть репарирован позже.
Репликация обеспечивает самокопирование генетического материала. При этом, благодаря принципу комплементарности, весьма высока точность сопоставления нуклеотидных последовательностей дочерней цепи к матричной ДНК. Кроме того, ДНК — достаточно химически инертное вещество, что обеспечивает ее большую стабильность по сравнению, например, с РНК. Однако этого мало, так как ДНК все же может повреждаться внешними воздействиями, также могут возникать ошибки на этапе репликации. Поэтому в клетках должны существовать механизмы исправления повреждений и ошибок синтеза, т. е. выполняться репарация ДНК.
Существует целый ряд репарационных механизмов, выполняющихся на различных этапах синтеза ДНК, а также в зависимости от типа возникающих ошибок.
Все вместе репарационные механизмы существенно снижают частоту ошибок в молекулах ДНК и направлены на поддержание стабильности наследственного материала. Однако, поскольку не все изменения структуры ДНК устраняются, возникают мутации, благодаря которым на Земле возникло разнообразие живых организмов.
Устранение ошибок ДНК-полимеразой
Прежде всего сама ДНК-полимераза при наращивании новой цепи ДНК проверяет, тот ли нуклеотид присоединяется к растущей нити.
Существуют измененные формы азотистых оснований, которые могут комплементарно связываться с нуклеотидами матрицы. Так измененная форма цитозина может связаться с аденином. Полимераза присоединит этот конечный нуклеотид к растущей цепи, но он быстро перейдет в свою обычную форму — станет обычным цитозином. При этом водородные связи разрушаются (т. к. нарушается комплементарность), и на конце получается неспаренный нуклеотид, однако ковалентно соединенный с синтезируемой цепью. Полимераза не может далее наращивать цепь. Сама полимераза или связанный с ней фермент редактирующая эндонуклеаза отщепляют последний «неправильный» нуклеотид.
В результате такого механизма самокоррекции частота ошибок репликации снижается в 10 раз. Если присоединение ошибочного нуклеотида на этапе синтеза ДНК составляет 10-5, то репарационная активность полимеразы снижает их количество до 10-6.
Репарационные механизмы
ДНК-полимераза исправляет часть ошибок репликации, но не все. Кроме того, изменения в последовательности нуклеотидов ДНК возникают и после ее удвоения. Так могут теряться пуриновые основания (аденин и гуанин), дезаминироваться цитозин, превращаясь в урацил. Эти и другие изменения возникают обычно из-за содержащихся в окружающей хромосомы среде определенные химически активных вещества. Ряд подобных соединений нарушает нормальное спаривание оснований. Под действием ультрафиолетового излучения два соседних остатка тимина могут образовать связи между собой, возникают тиминовые димеры.
Существует прямая репарация, когда, если это возможно, ферментативно восстанавливается исходная структура нуклеотидов, без их вырезания.
Эксцизионная репарация
Эксцизионная, или дорепликативная, репарация осуществляется до очередного цикла репликации.
Существует класс ферментов, обнаруживающих измененные последовательности нуклеотидов в одной из комплементарных цепей ДНК. После этого происходит удаление ошибочного участка и его замена вновь синтезированным. При этом матрицей служит участок комплементарной «правильной» нити.
Ферменты репарации обычно обнаруживают ошибки на новой нити ДНК, а не матричной. Между двумя цепями одной молекулы ДНК небольшое различие, заключающееся в степени метилирования азотистых оснований. У дочерней цепи оно отстает от синтеза. Ферменты распознают такую цепь и именно на ней исправляют участки, которые так или иначе не комплементарны участкам старой цепи. Кроме того, сигналами могут служить разрывы нити, которая у эукариот синтезируется фрагментами.
Фермент эндонуклеаза способна обнаруживать утрату пуриновых оснований. Данный фермент разрывает фосфоэфирную связь в месте повреждения. Далее действует фермент экзонуклеаза, который удаляет участок, содержащий ошибку. После этого дыра застраивается согласно комплементарности матрице.
ДНК-гликозилазы – целый класс ферментов, распознающих повреждения ДНК в результате дезаминирования, алкилирования и других структурных изменений ее оснований. Гликозилазы удаляют именно основания, а не нуклеотиды. После этого участки нити ДНК без оснований репарируются также как при «починке» пуринов.
Следует отметить, что дезаминирование азотистых оснований может привести к невозможности восстановления исходной последовательности нуклеотидов. Происходит замена одних пар оснований другими (например, Ц-Г заменится на Т-А).
Ферменты, удаляющие участки с тиминовыми димерами, распознают не отдельные ошибочные основания, а более протяженные участки измененной ДНК. Здесь также происходит удаление участка и синтез на его месте нового. Кроме того димеры тимина могут устраняться самопроизвольно под действием света — так называемая световая репарация.
Пострепликативная репарация
Если дорепликативная репарация не исправила измененные участки ДНК, то в ходе репликации происходит их фиксация.
266. Репарация днк. Виды репарации.
Одна из дочерних молекул ДНК будет содержать изменения в обоих своих нитях. В ней одни пары комплементарных нуклеотидов заменены на другие, или появляются бреши во вновь синтезированной цепи напротив измененных участков матричной.
Система пострепликативной репарации способна распознавать такие изменения ДНК. На этом этапе устранение повреждений ДНК осуществляется путем обмена фрагментами (т. е. рекомбинацией) между двумя новыми молекулами ДНК, одна из которых содержит повреждение, другая — нет.
Так происходит с димерами тимина, которые не были удалены на предыдущих этапах. Между двумя рядом стоящими тиминами присутствуют ковалентные связи. Из-за этого они не способны связываться водородными связями с ковалентной цепью. В результате, когда на матричной цепи, содержащей тиминовый димер, синтезируется дочерняя цепь, в ней образуется брешь. Этот разрыв распознается ферментами репарации. Понятно, что правильного участка у данной молекулы ДНК нет (одна нить содержит тиминовый димер, другая — дыру). Поэтому единственный выход — это взять участок ДНК со «здоровой» молекулы, который берется с матричной цепи этой молекулы ДНК. Образующаяся здесь дыра заполняется по принципу комплиментарности.
SOS-система
Значительная часть повреждений ДНК устраняется с помощью описанных репарационных механизмов. Однако если ошибок остается слишком много, то обычно включается так называемая SOS-система, состоящая из своей группы ферментов, которые могут заполнять дыры, не обязательно соблюдая принцип комплементарности. Поэтому срабатывание SOS-системы часто служит причиной возникновения мутаций.
Если же изменение ДНК слишком существенное, то репликация блокируется, и клетка не будет делиться.
ДНК каждой клетки человека теряет за сутки около 5000 пуриновых остатков вследствие разрыва N-гликозидной связи между пурином и дезоксирибозой (рис. 4-22). Тогда в молекуле ДНК на месте этих оснований образуется участок, лишённый азотистых оснований, названный АП-сайтом (AP-site, или апуриновый сайт). Термин "АП-сайт" используют также в тех случаях, когда из ДНК выпадают пиримидиновые основания и образуются апиримидиновые сайты (от англ, apurinic-apyrimidinic site). Этот тип повреждений устраняет фермент ДНК-инсертаза(от англ, insert — вставлять), который может присоединять к дезоксирибозе основание в соответствии с правилом компле-ментарности. В этом случае нет необходимости разрезать цепь ДНК, вырезать неправильный нуклеотид и репарировать разрыв.
Дезаминирование
Реакции дезаминирования цитозина и превращение его в урацил (рис. 4-23), аденина в гипоксантин, гуанина в ксантин происходят значительно реже, чем депуринизация, и составляют 10 реакций на один геном в сутки. Исправление этого вида спонтанного повреждения происходит в 5 этапов (рис. 4-24). В репарации принимает участие ДНК-N-гликозилаза,гидролизующая связи между аномальным основанием и дезоксирибозой (первый этап), в результате образуется АП-сайт, который распознаёт фермент АП-эндонуклеаза (второй этап). Как только в цепи ДНК возникает разрыв, в работу вступает ещё один

Рис. 4-21. Система репарации ошибок репликации.
1 — белок mut S "узнаёт" некомплементарную пару и присоединяется в этом участке ДНК; 2 — белки mut H взаимодействуют с метилированной по аденину последовательностью материнской цепи -GATC-; завершается формирование ферментативного комплекса после присоединения mut L; 3 — комплекс определяет вновь синтезированную цепь по отсутствию метилированного остатка аденина в последовательности -GATC- и разрывает её; 4 — экзонуклеаза удаляет фрагмент дочерней цепи ДНК, содержащий ошибку; 5 — ДНК-полимераза β по принципу комп-лементарности застраивает брешь; 6 — ДНК-лигаза З’-конец вновь синтезированного фрагмента соединяет с основной цепью и завершает репарацию ошибки.
фермент — АП-экзонуклеаза, который отщепляет от цепи дезоксирибозу, лишённую основания (третий этап). В цепи ДНК появляется брешь размером в один нуклеотид. Следующий фермент ДНК-полимераза р к З’-концу разорванной цепи присоединяет нуклеотид по принципу комплементарности (четвёртый этап). Чтобы соединить два свободных конца (3′-конец встроенного нуклеотида и 5′-конец основной цепи), требуется ещё один фермент — ДНК-лигаза (пятый этап). Нерепарйруемо и поэтому опасно дезаминирование метилированного цитозина. Продукт его спонтанного дезамжнирования — тимин,

Рис. 4-22. Депуринизация — спонтанное удаление аденина или гуанина.

Рис. 4-23. Продукты спонтанного дезаминирования различных оснований ДНК. Все продукты дезаминирования (урацил, гипоксантин, ксантин) нехарактерны для состава ДНК и поэтому довольно легко распознаются ферментами репарации.
нормальное для ДНК основание, которое не распознаётся ДНК-N-гликозилазой.
Б. Индуцируемые повреждения
Индуцируемые повреждения возникают в ДНК в результате воздействия разнообразных мутагенных факторов как радиационной, так и химической природы.
Образование димеров пиримидиновых оснований
Под действием УФО двойная связь между С5 и С6 атомами углерода в составе пиримидиновых оснований (тимине и цитозине) может разрываться. Атомы углерода остаются связанными одной связью. Расстояние между параллельными плоскостями оснований полинуклеотидной цепи, в которых произошёл разрыв., равно примерно 3,4  . Это расстояние позволяет освободившимся валентностям между С-С атомами пиримидиновых оснований, расположенных последовательно в цепи ДНК, сформировать циклобутановое кольцо (рис. 4-25). В зависимости от того, какие основания соединены в димер, их называют димерами тимина, цитозина или ти-мин-цитозиновыми димерами. Удаление пиримидиновых димеров происходит под действием фотолиазыФермент расщепляет вновь образовавшиеся связи между соседними пиримидиновыми основаниями и восстанавливает нативную структуру.
. Это расстояние позволяет освободившимся валентностям между С-С атомами пиримидиновых оснований, расположенных последовательно в цепи ДНК, сформировать циклобутановое кольцо (рис. 4-25). В зависимости от того, какие основания соединены в димер, их называют димерами тимина, цитозина или ти-мин-цитозиновыми димерами. Удаление пиримидиновых димеров происходит под действием фотолиазыФермент расщепляет вновь образовавшиеся связи между соседними пиримидиновыми основаниями и восстанавливает нативную структуру.
Репарация ДНК
В фотолиазе есть участок, либо сам поглощающий фотоны (в синей части спектра), либо связывающийся с кофакторами, адсорбирующими свет. Таким образом, свет активирует фотолиазу, которая распознаёт димеры в облучённой ДНК, присоединяется к ним и разрывает возникшие между пиримидиновыми кольцами

Рис. 4-24. Репарация АП-сайтов с участием ДНК-М-гликозилазы и АП-экзонуклеазы.
связи. После этого фермент отделяется от ДНК.
252-253


Молекулярная генетика
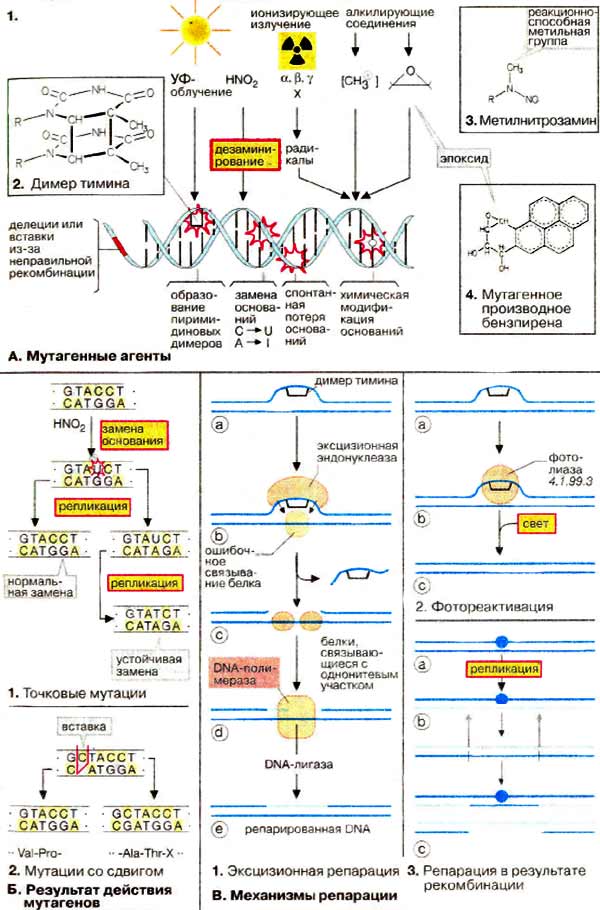 Генетическая информация кодируется последовательностью оснований ДНК и поэтому изменения в структуре или последовательности азотистых оснований приводят к мутациям. Многие мутагены вызывают нарушения регуляции роста и деления клеток и поэтому являются канцерогенными. Изменение в структуре генов (мутация) — важный фактор биологической эволюции. В то же время слишком высокая скорость мутаций ставит под вопрос существование индивидуальных организмов или целых видов. Поэтому клетки обладают механизмами восстановления (репарации), которые корректируют большинство изменений ДНК, вызываемых мутациями.
Генетическая информация кодируется последовательностью оснований ДНК и поэтому изменения в структуре или последовательности азотистых оснований приводят к мутациям. Многие мутагены вызывают нарушения регуляции роста и деления клеток и поэтому являются канцерогенными. Изменение в структуре генов (мутация) — важный фактор биологической эволюции. В то же время слишком высокая скорость мутаций ставит под вопрос существование индивидуальных организмов или целых видов. Поэтому клетки обладают механизмами восстановления (репарации), которые корректируют большинство изменений ДНК, вызываемых мутациями.
Мутации могут возникать в результате либо воздействия физических или химических факторов, либо ошибок в процессе репликации и рекомбинации ДНК.
Одним из наиболее важных физических мутагенов является ионизирующая радиация. Она приводит к образованию в клетке свободных радикалов (молекул с неспаренными электронами, см. с. 20), которые исключительно реакционноспособны и могут повредить ДНК. Коротковолновый ультрафиолетовый свет (УФ) также оказывает мутагенное действие, особенно на клетки кожи. Наиболее распространенным химическим изменением, вызванным ультрафиолетовым облучением, является образование тиминовых димеров (2). когда два соседних тиминовых оснований ковалентно связываются друг с другом. Это приводит к ошибкам при считывании ДНК во время репликации и транскрипции.
Из множества химических мутагенов здесь приведен только несколько примеров. Азотистая кислота (HNO2) и гидроксиламин (NH2OH) дезаминируют азотистые основания, т. е.
превращают цитозин в урацил, а аденин в инозин. Алкилирующие соединения имеют реакционные группы, которые могут образовывать ковалентные связи с азотистыми основаниями, входящими в ДНК (см. с. 388). Метилнитрозамины (3) распадаются с образованием реакционноспособного метилкатиона (СН3-), который метилирует группы ОН и NH2 в ДНК. Ароматический углеводород бензпирен сам по себе безвреден, но в результате метаболической трансформации образует производные, обладающие канцерогенным действием (4). За счет гидроксилирования одного из колец он превращается в реакционноспособный эпоксид, который алкилирует аминогруппу гуанина и других азотистых оснований. Токсичен и свободный радикал бензпирена.
Азотистая кислота вызывает точковые мутации (1). Например, С превращается в U, который в следующем цикле репликации образует пару с А (вместо G). после чего изменение принимает необратимый характер. Мутации типа вставки или выпадения некоторого числа нуклеотидов, не кратного трем, ведут к ошибочной трансляции всей ДНК, поскольку они сдвигают рамку считывания (2).
При трансляции измененная мРНК будет интерпретироваться рибосомами по-другому, приводя к совершенно иной аминокислотной последовательности, что показано на простом примере (2).
Важным механизмом удаления повреждений в ДНК является эксцизионная репарация (1). Специфическая нуклеаза удаляет небольшой сегмент ДНК, включающий поврежденный участок. Удаленный участок восстанавливается ДНК-полимеразой, использующей в качестве матрицы комплементарную цепь. Наконец, оставшийся одноцепочечный разрыв закрывается ДНК-лигазой. Тиминовые димеры могут быть удалены фотореактивацией (2). Специфическая фотолиаза связывается с дефектным участком ДНК и после облучения расщепляет димер с образованием отдельных нуклеиновых оснований. Третий механизм — это репарация в результате рекомбинации (3, показано в упрощенном виде). В этом процессе участок, содержащий повреждение, пропускается во время репликации. Образующаяся брешь закрывается путем сдвига соответствующего сегмента из правильно реплицированной второй цепи. Новая брешь ликвидируется с участием полимераз и ДНК-лигаз. В завершение первоначальный дефект (1) устраняется путем вырезания.